As I mentioned yesterday, Malcolm Hughes and/or the University of Arizona Laboratory for Tree-Ring Research had gone to the trouble of blocking my IP address from accessing their website. Thanks to the help of a couple of CA readers, I was able to circumvent the block. This is the sort of petty behavior that gives (or should give) the Team a bad name in a couple of senses – it both makes them look bad and it’s pointless because it’s readily circumvented. Given the relative ease of circumvention, you’d think that they wouldn’t bother, but they don’t seem to care. One CA reader wrote and asked me if he could locate what I was looking for. Originally I was just going to check their listserv, which I did, and there was nothing new there. But since I’d been blocked, I went to a little extra trouble to check the site and it was worth it.
As I’ve mentioned previously, the Sheep Mountain bristlecones were re-sampled in 2002 but nothing has been published about it. Based on my experience with mining promotions – delayed results nearly always are bad as promoters find way of ensuring that good results are released on time or early, I’ve predicted that the Sheep Mountain update would not show elevated growth rates. (Yeah, there will be a “good” reason why any results took more than 5 years to be published, but “good” results would have been accelerated so that they were available for AR4. My bet offer was discussed by James Annan here.
An LTRR PhD student, Linah Ababneh, has been working on bristlecones (she got their “Bristlecone Prize” in 2002). I’ve emailed her in the past without any response and, from time to time, I google or otherwise check for information on her work. In yesterday’s browse through the LTRR website, I checked the page for theses and, sure enough, Linah Ababneh’s thesis was now online. It’s a must-read for CA readers.
Her starting point is Graybill and Idso 1993 and their reported difference between chronologies for whole-bark and strip-bark trees. She sampled 100 bristlecones – 50 at Sheep Mountain and 50 at nearby Patriarch Grove, with 25 whole-bark and 25 strip-bark – for each site. She has an overhead photo showing the location of each site. In addition to making a composite chronology, she made 4 chronologies for the bark x site combinations. She said that the chronologies extended from 1071-2003 and would be archived at ITRDB “after December 2006”; they aren’t.
Before I discuss how she reported her findings, let me simply show two illustrations of her chronologies. The first figure shows all 4 chronologies from 1600 to 2003 (with an 8 year smooth). BTW her thesis has 3 chapters which are drafted in journal style – two of the chapters are aimed at a dendro audience and a third is aimed at an archaeological audience (as she associates information from tree rings with settlements in the White Mountains.) In the two dendro chapters, she only illustrates her chronology for the 1600 on period. Seems odd, given the interest in millennial chronologies. Anyway here is the composite. Look at the right hand side of the chronology for the precipitous decline in Sheep Mountain ring widths in the past 20 years – which affected both strip-bark and whole-bark chronologies.
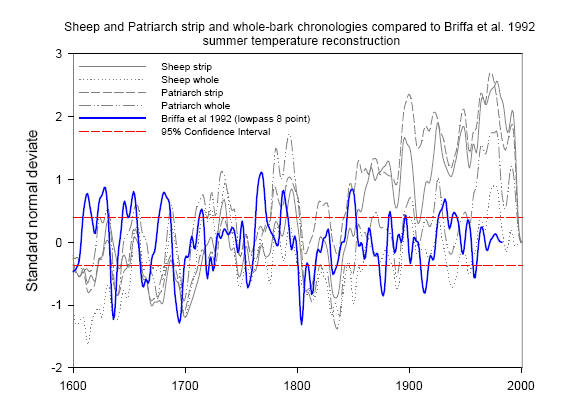
Abaneh Figure 3.Sheep and Patriarch strip and whole bark tree-ring width chronologies compared with Briffa et al. 1992 summer reconstruction based on tree-ring density. All chronologies are filtered after converting to z-scores to match the scale. Tree-ring data and Briffa et al. (1992) data are filtered using an 8-year low-pass filter (Fritts 1976). Confidence Intervals are based on the Briffa et al. (1992) series.
In her chapter linking to archaeology (scheduled for submission to Quaternary International), she has the following illustration of her long composite chronology (including strip bark), which she compares to Lamarche 1974 (but not Graybill’s chronology used in the MBH PC1: note- I’ll plot this up later or you can see Graybill’s Sheep Mountain chronology in the MM articles.) As you see, for whatever reason, her chronology differs materially from the Lamarche chronology (and even more from Graybill.)
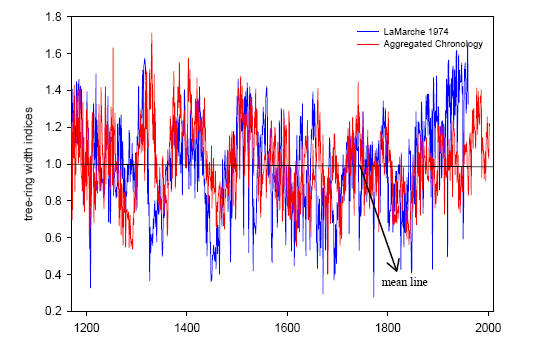
Ababneh chapter 3 Fig.4. LaMarche (1974) overlaid by Aggregated chronology from Strip-bark and whole-bark of Patriarch Grove and Sheep Mountain develop in this study for visual comparisons. Both chronologies are tree-ring width indices.
Here is the same chronology plotted a little differently. IF you look closely, you can see that this is the same as the red series in Figure 4.

Ababneh Fig. 5. Cold and warm periods as inferred from tree ring widths chronology (Ababneh, 2006, This study) fluctuations above and below the mean after normalizing, whole-bark and strip-bark chronologies are grouped together from two sites Patriarch Grove and Sheep Mountain.
For comparison, I’ve plotted up the archived Lamarche (ca046) and Graybill (ca534) chronologies for Sheep Mountain as shown below. The Lamarche chronology ends in 1970; the Graybill chronology goes to 1987, but Mann only uses it to 1980. There’s something inconsistent between the Lamarche version illustrated by Abaneh and the archived version: they look very similar, but the archived version is about 20% higher (it touches 1.8 while the Ababneh illustrated version only goes to 1.6). The archived Graybill version goes as high as 2.4 – why wouldn’t Ababneh have illustrated and discussed it. I’ve marked the 1980 value of the Graybill Sheep Mountain series, as this is the last year used in Mann’s PC1. I’ve plotted the values from 1980-1987 in grey and one sees an incipient divergence problem. But the more interesting issue is that Ababneh’s chronology isn’t just calling into question the Divergence Problem; it’s calling into question the reality of the Lamarchel growth pulse – and even more dramatically, the Graybill growth pulse (which is never discussed.)
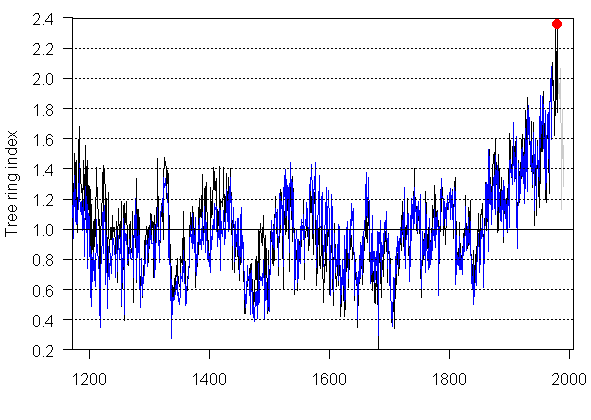
Plots of Archived Lamerche (ca046 -blue) and Graybill (c1534 – black/grey) chronologies for Sheep Mountain.
Her dissertation committee included Malcolm Hughes and Jeffrey Dean, but not a whisper about the “Divergence Problem”. She reported that she had confirmed a significant difference between strip-bark and whole-bark chronologies, but was unable to explain the difference.
I noted that, at Almagre, measurement data is missing for some Almagre trees where tags have been located, but measurements have not been archived. Ababneh made the following interesting comment about sampling differences:
As shown in Figure 13 (5 samples ), whole-bark trees from Patriarch Grove seem to have a mixed form of growth, i.e. three trees have an increase in tree-ring width, while two do not show such an increase. In comparison, Figure 8 (18 samples) illustrates that none of the whole-bark trees have an increase in tree-ring widths over the period shown. This shows how inadequate sampling might lead to different conclusions.
In chapter 2, Ababneh assessed climatic relationships between these chronologies, finding little relationship. She states:
Because the results of this study do not provide enough evidence to support the hypothesis that whole-bark tree growth is controlled by temperature, a temperature reconstruction cannot be established based on the chronologies and analyses developed in this study….
Until these limitations [other factors limiting growth] are taken into consideration when modeling tree growth, and until further measurements are obtained that involve a longer instrumental climate record from the same elevation of the research sites, the positive or negative effects of temperature cannot be substantiated.
The following paragraph is amusing. After re-iterating the evidence against a temperature signal in these chronologies one more time, she cites IPCC 2011, MBH99 and Mann et al 2000 as “a new and growing body of literature … that upper forest border trees respond …to the observed global temperature increase”:
The results reported in this paper are partially in accordance with other studies that support the hypothesis of a moisture related signal in bristlecone pine. Wright and Mooney (1965) concluded that bristlecone pine responds to a precipitation gradient rather than to cool alpine temperatures. Similar results were reported from a three year ecophysiological study of bristlecone pine by Fritts (1969). Such findings are not unexpected since the White Mountains are xeric with annual precipitation not exceeding 41.5 cm. Graybill and Idso (1993) and Graybill and Funkhouser (1999) compared tree-ring widths from the western United States, including one of the sites investigated in this study (Patriarch Grove), and found similar results: a low correlation with temperature that prevents use for temperature reconstruction, a negative correlation with the previous year’s temperature, and a highly significant tree growth response to spring precipitation. Bunn et al. (2003) and Tang et al. (1999) implicated soil moisture sensitivity especially in the strip-bark trees. However, a new and growing body of literature supports the other part of the results provided within; that is, upper forest border trees respond differently to climate variability including the observed global temperature increase (IPCC 2001; Mann et al. 1999; Mann et al. 2000), and a growing season temperature signal can be detected on decadal time scales (Hughes and Funkhouser 2003).





79 Comments
As I’ve been repeating, onging, here, for 2 years:
“Wright and Mooney (1965) concluded that bristlecone pine responds to a precipitation gradient rather than to cool alpine temperatures. Similar results were reported from a three year ecophysiological study of bristlecone pine by Fritts (1969). Such findings are not unexpected since the White Mountains are xeric with annual precipitation not exceeding 41.5 cm. Graybill and Idso (1993) and Graybill and Funkhouser (1999) compared tree-ring widths from the western United States, including one of the sites investigated in this study (Patriarch Grove), and found similar results: a low correlation with temperature that prevents use for temperature reconstruction, a negative correlation with the previous year’s temperature, and a highly significant tree growth response to spring precipitation. Bunn et al. (2003) and Tang et al. (1999) implicated soil moisture sensitivity especially in the strip-bark trees.”
[NOTE: The White Mountains alluded to here are the ones in Inyo County, California, not the ones in New Hampshire]
I noticed she cited the IPCC 2001 in the last sentence above but doesn’t seem to list any portion of the IPCC 2001 report in her cited reference section. I’d be curious to see the context of the IPCC’s statement on the matter. If it’s simply a reference cited by the IPCC itself, it has no business being a cited reference here (original reference should have been read and cited instead). So it would appear the lone “substantiated” support is Mann et al (x2). Why isn’t anybody else contributing to this “new and growing body of literature?”
So someone DID update the proxies.
Now we wait for the “peer review” to start.
Wanna bet that Graybill, Idso, and Funkhouser are on the “peer list”?
Post Barton (2005), I think any school, and by policy its graduate students, should be financially accountable for delivery of promised or required data. Particularly since this involves public policy issues and presumably public funding. I’m not sure whether to call this missing data (it’s after 2006…) for a recent PhD, a PhD for noncompleted work, or a form of academic / scientifc misconduct. It seems hard to justify not doing some kind of cut and paste of at least raw data in a text or .xls file with some kind of headers after the effort of a 145 page thesis unless one is worried that the data will be shown to compromise/contradict the claimed results. In fact such a required filing *with the thesis* would seem an excellent opportunity for a true *public* defense (open source review) to identify weak or flawed theses.
She gambled and lost. Her thesis will forever be a non-entity.
#4. I’d be surprised if Linah Ababneh is to blame. IF Hughes et al are going to the trouble of blocking me from their website, I’m sure that they keep an eye on what’s archived. The issue is Hughes and Dean, not Ababneh.
#3. I’ll bet big money that Graybill is not on the peer review list.
BTW this thesis has been “peer reviewed” since it’s been accepted as a PhD Thesis. It’s been reviewed by Hughes, Dean and two others. I’m sure that PhD theses are reviewed more competently than an average journal article.
To get an IP block at a university server, you’d have to have the university IT people in on it. I seriously doubt that the lab would have it’s own dedicated server. I wonder what their IT services people have to say about this? And the university administration? What possible pretext could they have for blocking IP addresses? I don’t know how to raise H E double hockeystick about this, but someone with a connection to the university might. There can’t be a reasonable explanation.
Just as a side note, if you ever get your IP blocked for whatever reason, simply go into Google translator, or babelfish and translate the page from any language back into English. Because the web page is in English already you’ll simply see the web page through the translators proxy. Easy and quick.
8, I think Steve’s already found a proxy, but the question remains, what’s up with the banana republic tactics? Is this any way for a professor of science to behave?
or use one of the hundreds, if not thousands of free anonymous proxies available on the net, such as http://www.pimpmyip.org
They can’t block you really, but it’s very telling they are willing to go to such lenghts. It’s childlish and makes them look bad, nothing else.
Since the manuscripts that constitute her thesis are not published yet, it is scarcely unreasonable that the data are not yet archived.
#11. Relative to the archiving failures that are endemic in the trade, I agree that this is a very slight non-archiving case – although the thesis did say that it would be archived “after December 2006” so shw should have done what she said. Given that Esper’s never archived any measurement data and Thompson’s sample data remains unarchived – in some cases, after 20 years -, it’s small potatoes.
re: #12
But Steve, she said it would be archived sometime after December 2006 so whenever she does indeed archive it she will have done what she said. Sure, some “unreasonable” people may think she meant that it would already be archived as of Jan 1, 2007, but the key point is what is meant by “would be”, which is a varient of the verb “is” and as we all know, knowing what the meaning of the word “is” is is serious business.
BTW, don’t you think my using the phrase ‘”is” is is’ is clever?
Steve
your getting to close to the truth and their getting nervous…don’t stop
many, many thanks
Steve
sorry, that would be “your getting too close…..”
No, it should be “you’re getting too close….”
and “they’re getting nervous”
I wouldn’t over-rate my impact on them. They’re probably just being annoying and trying to make me waste time on little things.
But what does that say about their priorities?
It says one of their priorities is “Let’s mess with Steve McIntyre.”
😀
Wow, this is a tough crowd. Can’t a student throw in a bit of context of the scientific climate of her day? Shouldn’t she?
Seriously, you get so used to this kind of monkey business, that you need to take a couple steps back, and ask if this is how a professional scientist should be behaving (never mind the IT help who went along with it, and presumably university administrative staff who at the very least looked the other way). Really, what is going on?
However, a new and growing body of literature supports the other part of the results provided within; that is, upper forest border trees respond differently to climate variability including the observed global temperature increase (IPCC 2001; Mann et al. 1999; Mann et al. 2000), and a growing season temperature signal can be detected on decadal time scales (Hughes and Funkhouser 2003).
Please forgive my ignorance: I am very much a novice with these things, but is this an example of that magical “effect at a distance” wherein the local conditions don’t affect the readings but the “global” readings do? What’s that called?
As for who is blocking your IP addy, it can be no other than Mr. Al “I’m winning all the prizes to make up for losing Florida” Gore. He invented IP, after all, and has a Master Console from which he can manage all of the switches, routers, and firewalls that connect thereto.
“To get an IP block at a university server, youd have to have the university IT people in on it. ”
That by another Larry.
This is something I can say something about with some authority.
The other Larry is mistaken. There are several ways a distant IP address can be blocked, many of them on the machine where the files are resident (see “IP tables” and “firewall” for entry points to discussions) or server-local routers and WiFi’s.
Who knew that there is a Bristlecone Chapter of the California Native Plant Society CNPS and its seem that in 2002 they make a grant to Ababnah to study the response of BCP and Foxtail Pines to CO2 and Nitrogen.
Steve M
This was clearly an unexpected find. A somewhat embarrassing one at that. It does beg the question what other ‘bad news’ for hockey stick manufacturers exists out there that is yet to be discovered? Keep up the good work Steve. as I’ve posted before, it really is now only a matter of time before all this make sit into the MSM. The tide is most definitely turning. As each day passes I’m getting more and more friend sof mine who were previously susceptible to the propaganda put out by the BBC now becoming increasingly sceptical. So much so that i’m having to remind them to look at the evidence themselves (as I have done) and to make up their own minds and certainly not to rely on what they are fed by the MSM.
KevinUK
I don’t think anyone should be criticising the student here. As far as I can see she has gone out, collected the data, analysed it and presented it. Of course her conclusions are going to be vague, since the results (These results confirm that water is the essence of life in the desert) undermine some precious ideas about “treemometers”.
Look at her examiners and supervisors- she’s got to play safe.
Are Hughes and Dean getting any federal funding for their work? If they are, and they are pulling these kind of shenanigans and their university stands behind them, then their funding should be pulled. This is getting ridiculous — blocking sites, not archiving data. We’ve got a $20 trillion climate problem and a bunch of professors on the public teat playing games — with my damn money! If they can’t publically share and archive data, then what the hell am I paying for? What is our government getting? This is serious — we are talking about about annual climate controls that will cost the average family the equivalent of a Harvard education to implement and they play silly, stupid games. To hell with them. Enough is enough.
24, You think these guys are capable of that? And if they’re doing all of that (which takes hours for a competent IT guy), what aren’t they doing?
As an ex IT guy, I’ll tell you it is a no-brainer to block IP-addresses on a Linux box, any of a variety of firewalls, or any of a variety of routers, all of which can be local to the server.
I am under NDA as to why I was fired from the University, but even when I was there there were folks all over campus who were better skilled than the average “IT guy”.
30, Ok, but do you think that: 1) the university allows them to webhost on their own departmental box (rather than on the university server outside of the router), and 2) these climate guys can figure out how to run Apache? They still use Fortran, for gawd’s sake.
My sympathies are with Ms Ababneh. She had to put two statements contradicting each other into her dissertation to both tell the truth and placate her professors. When I did my MA thesis I found I had to exclude certain conclusions to maintain minimal civil relations with my primary professor. That is, he had clearly mixed up two samples when he did his analysis, and when I tried to tell him that as politely as possible he began screaming “They were right when they told me you hated me!” (Long story like a bad soap opera.) Some researchers deal with their data as though they were their own children, to be treasured and defended.[snip]
Furthermore (and here I am off the edge of what I know well), it seems to me that the web server software (“Apache”, “IIS” e.g.) allows filtering within the file structure or in the server configuration.
I am not knowledgeable about either Apache or IIS.
#31 You’re doing yourself a disservice by underestimating the intelligence of professors. A lot of them that I’ve met know newer languages, but use Fortran for its speed and documentation (and the ease of use of pre-written codes).
Plus it’s pretty common for various internal groups to run their own servers. Hell, up until recently, each department ran their own server at my university (e.g. physics.queensu.ca, me.queensu.ca, etc.)
#34. My guess is that LTRR has their own server and that the blocking was done directly.
I could be wrong. I may be confusing universities with corporations. I know no corporation would ever let departments run their own webservers. At least no corporation I’ve ever been near. The pecking order may be different at universities.
Abaneh is at least not alone. Olga Solomina had similar results. Check out figure C.
Re IP Blocking.
Steve, try telneting to the web server on port 80 and according to the response you should be able to tell roughly how they are blocking you.
“Connection refused” Blocking you with a firewall in a nice manner, ie. they respond to your request with a refusal.
“Connection timed out” Blocking you with a firewall in a nasty manner. ie. they simply drop your request packet and don’t tell you about it. This can take a minute or so.
“Connected” followed by “Connection closed” The web server software is rejecting your connection. No firewall involved.
If you do try this make sure you telnet to port 80 otherwise they might accuse you of trying to hack the server.
38, I think you may have hit the answer. They assumed that Steve hacked Mann to get the “censored” directory (it was lying there in broad daylight). Of course if Steve were a super hacker, their blocking attempts would be futile, anyway. It’s so silly.
Larry (no 36),
If a Department has enough money to buy a webserver, then the Department is going to run it.
Professors with enough grant money have been buying computers for their own research for decades.
Roger Bell
The graphs in Ababneh’s thesis and Appendices made me wonder how this relates to the solar irradiance. So I graphed the solar reconstruction data (Lean, 2004) to compare.
Here is Ababneh Figure 3 with the Lean solar in red.
To get a better idea of the correlation of the cycles, here is a close up of the more recent portion of the same figure.
Ababneh also provides a climate reconstruction graph in terms of temperature and precipitation. Here it is from 1600 on with the Lean solar in red.
Steve,
Are you sure your IP address was blocked and that this isn’t just a case of over-aggressive software on the part of their IT department? Between spam, porn, and file-sharing (i.e. music), it wouldn’t be surprising if servers block IP addresses for oddball reasons. Maybe their software is tagging “climateaudit” as an acronym for “climax” or “audio”. It might sound odd, but I’m inclined to suspect a mundane explanation like this over a conspiracy of sorts.
40, not to beat this to death, but it’s not a question of budget, it’s a question of organization. Businesses don’t do it that way. They keep webserving locked up on a server outside of the corporate router/firewall. They don’t want every Tom, Dick, and Harry from Russia and wherever else running around on the inside of the local network. Universities must be different.
42, that’s also possible. I’ve seen some pretty innocuous things caught in my spam filter as porn.
I cannot find a tree ring graph in AR4, but tree rings play a role in it for the argument the last half of the 20th century was the warmest in the Northern Hemisphere in the last 1300 years. See pp. 702-703,
In other words, tree rings are still critical for IPCC arguments about anthropogenic AGW.
#42
In order to do that the web server would have to do a reverse lookup on Steve’s IP address to resolve it as coming from climateaudit. In general, web servers don’t do that as it adds a delay to them serving the pages and increases network traffic. It also makes the web server performance dependent on a DNS server
Further on #45
AR4 ch. 6.6.1 expands on the tree rings. Ice cores are the other natural “thermometer” which I find even more problematic.
My last go at it–if you want to believe “IT guys” have special powers, go to it.
But at the university I was fired from, prior to my firing, ALL of the significant research data-bases were on departmental servers operated (in most cases, over the strenuous objections of most of the “IT guys”) by faculty members. “Operated by” included controlling access to the data on the servers.
Y’all really need to get a grip on what is actually real out there. You do not want to look like one of them with their narratives and nuances.
I don’t know if the IP addresses in question are blocked or not. I don’t know, if they are blocked, where they are blocked.
But I do know that access can be blocked easily by anybody bright enough to do all the other things you say they do, right on the machine housing the data.
I’m out.
Well, I lied–I forgot address one issue.
“In order to do that the web server would have to do a reverse lookup on Steves IP address to resolve it as coming from climateaudit”
That too is wrong. The “firewall code” or the “IP Tables code” on the machine housing the data knows what the IP address is in the packets it is supposed to respond-to.
#42. I got right in when I used an anonymizer access. There’s nothing connecting my IP address to cliamteaudit on its face. Finally, I’ve been blocked previously (and equally pointlessly) by Mann and Rutherford. In the Mann case, I pursued this with the university IT department and they said that they had no jurisdiction over Mann’s server. So in this particular case, I think that there are grounds for surmising intention.
I don’t know how to do the telnet, but the inquiry hangs up for a while and only fails after some time has passed.
Re#43:
Yes, Larry, universities tend to be pretty decentralized, with most assets purchased/managed on a lab by lab basis, often with additional more-or-less independent facilities managing high-dollar equipment that may be used by multiple investigators. The amount of central control a university-wide IT department has varies widely from institution to institution.
Re #42:
Rob, I’m guessing that Steve doesn’t surf the web from the actual climateaudit server, so I doubt that domain name has anything to do with Steve’s access problems. Whether the blocking was intentional or not, the search it prompted resulted in a nice result in any case.
Just for fun regarding the IP blocking, here is a cite from the University of Arizona’s Committee on Ethics and Commitment:
“The Committee on Ethics and Commitment shall receive reports from the Research Integrity Officer and deal with questions of misconduct in research, scholarship, or creative endeavor; conflict of commitment; and facilities misuse.
In its deliberations it will use the current versions of the University policies on research integrity, professional commitment and proper facilities use.”
I wonder if “proper facilities use” includes selectively blocking people from reviewing research.
Frank,
If the block is truly deliberate, I think the intent is more to give Steve the finger rather than to prevent him of accessing the site. Clearly, someone who wants access to the site can get around the block by using another account or asking a friend to access it … they must know this.
Denying Steve access is basically the equivalent of saying “you can’t play in my sandbox!” It is amazingly silly … which is why I expressed doubts earlier. This is well below the level that I expect intelligent adults to sink!
I asked the University of Arizona to look into it and see if selective blocking of researchers has taken place. I’m an alumni of the Arizona system and find it disturbing.
http://www.vpr.arizona.edu/contactus
When Feynman figured out a fatal flaw in top secret safes, they sent out a memeo: “Keep Feynman away from you safe.”
I wanted to get back to the thesis for a minute. Nobody has said it yet, so I will. I found Linah Ababnehs PhD dissertation a pleasure to read. Rather than reading it to refute, as clearly some of you have, I read it for enlightenment. After all, Steve said it was a necessary read for CA patrons and I agree. I learned a great deal. Moreover, I managed to zip through the narrative in about an hour and half. It is a fast read for the true wonk. That is code for well written. I found a couple of type-os that seems ubiquitous in todays science papers, but I blame the editor, not the author. The paper felt non-ideological as I read it, which is saying a lot, considering whom her supervisors are.
Aside from a couple throw away lines added to appease the generals, she has done some good research and stated authoritatively where her analysis ran counter to the prevailing wisdom. I think we should cut her some slack and keep an eye on her in the future. She is a good one.
Ian
55, you know what that combination was, don’t you? Every file cabinet used 271828. Gee, I wonder how a physicist came up with that number?
Finding the “censored” directory was even easier than that.
RE 42 & 50:
If you were really curious, you could nmap it:
http://insecure.org/nmap/
#37
Look at the second and third authors on this article, our old friends, D’Arrigo and Jacoby.
RE: #59 – those two were probably the origin of the paen to the IPCC.
Since people picked on poor dougp’s grammar in #’s 14-17 I’m going to flag KevinUK in #26 for perpetuating the misuse of “begs the question”. This phrase has a very specific meaning in logic. It refers to the logical fallacy “petitio principii”, in which the conclusion of an argument is implicitly or explicitly assumed in one of the premises.
Now there are plenty of circular arguments at RealClimate for which this phrase would be properly applied, but what I think Kevin means is “prompts the question” or “raises the question”.
It’s probably too late to stop this misuse of the phrase, as I hear it constantly in the popular press, but I have to try.
Otherwise I completely agree with Kevin’s remarks.
Re #61:
Delighted to see that someone else is disturbed by the ubiquitous misuse of the phrase “begs the question”. What they usually mean to say is “prompts the question”. But I’m afraid the misuse has already been accepted by the latest editions of the Merriam-Webster Collegiate Dictionary.
aurbo
Isn’t that the dictionary that Rex Stout’s fictional detective Nero Wolfe used to tear pages out of so he could throw them in the fire? All my Nero Wolfe paperbacks are in a box somewhere and are so old they’re probably too fragile to read so I can’t easily check.
On IP blocking. You can do it with a .htaccess file almost trivially if you have the necessary local file permissions. Works on IIS too.
A tutorial: http://www.freewebmasterhelp.com/tutorials/htaccess/
I wanted the question to make me a sandwich, so I begged the question.
I actually got into a debate with somebody that claimed such a circular argument was not in fact a fallacy. It is, but it is an informal one (and can have valid structure).
Mark
It’s just an infinitely recursive argument….
The street is wet, and that caused the rain which made the street wet, which caused the rain.
#68: Actually, this is kinda what happened in Texas all summer.
“What possible pretext could they have for blocking IP addresses?”
As I recall, the pretext they had last time was that the blocking was essentially automatic, because McIntyre was downloading a lot of data, and his downloads were undistinguishable from a denial of service (DoS) attack. That said, I believe the reason McIntyre had to download so much was the way they divided up the data into so many separate pages, when it could have been 1% as big if put into a single file in the 1MB range.
Mr. McIntyre:
1) Please correct me if I made any major errors above.
2) Did this most recent IP block follow upon such a large series of downloads as to make plausible such a claim (i.e., that your downloads looked like a DoS attack?)
Steve: I’ve never downloaded any data from them and hadn’t even visited the site recently.
He wasn’t downloading a lot of data, his automated script was going into directories listed in robots.txt and they didn’t know what it was. Once it got worked out, the folks there said if it acts like a crawler/robot/agent/script, same difference, going into robots.txt directories is going into robots.txt directories. And then they let him go back to what he was doing in the first place the same ways! .Nobody was denied service.
I can’t see how that could be the case here, it sounds like he couldn’t even get into the site inthe first place but I forget the specifics.
L Nettles #25 Link
Does the use of wells nearby the tree stand need to be taken into account?
Observation.
According to Ababneh:
Interestingly, Stockton 1986 (or 1984, if you go by the reference list) was never published:
The idea that was unpublishable in 1984 become fashionable around the time of MBH98.
Has anyone read the Stockton report?
No discussion of Ababneh Fig 5, p 121?
Check out the “temperature reconstruction” ~AD1330, ~AD1400. So warm, the 20th c. is not even close.
Only one passing reference to nonlinear growth response (to P,T,C,N). Little attention to interactions between growth factors (P*T*C*N), and the problem this would cause for the linear additive growth model.
THIS is a PhD thesis?
bender, the chronology in Figure 5 is the same as the (red) chronology in Figure 4. I’ve added it into the post for clarity, since I agree that it shows the point in your observation more clearly. Isn’t it amazing how the Graybill chronology (the heart of Mann’s PC1 and the Hockey Stick) got excised from history?
Ababneh Fig. 5. Cold and warm periods as inferred from tree ring widths chronology (Ababneh, 2006, This study) fluctuations above and below the mean after normalizing, whole-bark and strip-bark chronologies are grouped together from two sites Patriarch Grove and Sheep Mountain.
From the Ababneh thesis, we have what might appear as a contradiction of statements here, but perhaps not in the realm of dendroclimatology.
Exactly. She does an uncanny job avoiding the new (bcp growth has declined since Stockton & LaMarche in the 1970s-80s!) while rehashing the old (G=f(P,T,C,N) is a weak statistical model of bcp growth). “In the dessert, water is the essence of life”. Tell me something I didn’t know already. Like: “1998-2005 temperatures are ‘unprecedented’ in a milll-yun years”.
I loved the Abaneh quote:
It appears that the “File Drawer” problem hasn’t gotten the attention it deserves in the tree ring community …
w.
One Trackback
[…] of the bristlecone proxy records collected by Graybill have failed replication, as shown by the Ababneh Thesis. Not only that, but one of the authors of M2008 (Malcolm Hughes) had to have known that, because he […]