There appear to be some important sampling differences between subfossil and modern (living) cores.
Briffa et al [CD 1992, 114] provide the following caveat with respect to using the RCS method (global curve fitting) for Tornetrask:
“In applying this [RCS] method, each individual tree ring series should ideally start with the innermost ring (preferably near the base) of each tree. In practice, there is invariably some uncertainty in assigning the biological age of the rings. Here we have simply assumed that the first year of the sample data represents the first year of tree growth. This will result in a variable but generally small (though systematic) underestimation of the true ring age. In the case of the density data, the effect will be negligible. In the ring-width data, the final standardization curve probably slightly underestimates the width of young trees and could therefore impart a small positive bias to the standardized ring-width indices for young trees in a number of series. However, this effect will be insignificant when the biased indices are realigned according to calendar growth years and averaged with many other series.”
I have not so far located a comparable caveat in connection with the Polar Urals data set in Briffa et al 1995, 1996. Merely looking at plots of individual Polar Urals cores (see here for a complete set) shows a considerable difference between the incidence of juvenile growth in subfossil and modern cores. Nearly all modern cores have a significant juvenile growth pulse, while nearly all subfossil cores do not (core 862450, dated to 914-1130 but whose date is really uncertain, is a notable exception.) This certainly suggests that there is some sort of sampling bias between living and dead cores – which may affect the validity of the assumption that the first preserved ring of a subfossil core should be given a year 1 age.
This impression is somewhat codified in Figure 1 below (following a Briffa et al 1996 format) showing the mean ring width by 5-year age groups for modern (start >1635) in solid black and subfossil (start<1635) in dashed red for RW (top) and MXD (bottom).
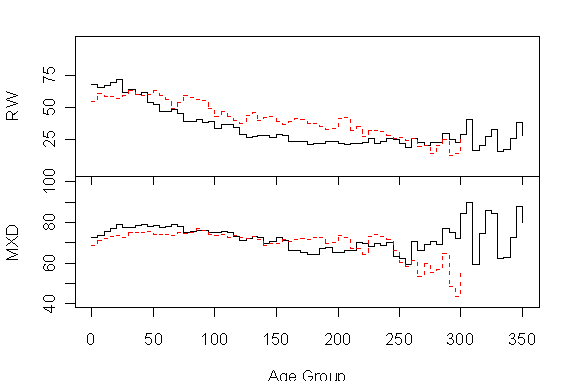
Figure 1. Polar Urals. RW (top) and MXD (bottom) for modern (solid black) and subfossil (dashed red).
For comparison, Figure 2 shows the scan of Figure 3 from Briffa et al , NATO 1996 — the bottom panel is Polar Urals RW and the third panel is Polar Urals MXD. The labels show parameters for the fitted curves.

Figure 2. Scan of Figure 3, Briffa et al [1996]. Original caption: “Regional Curve Standardization (RCS) functions for the Swedish and Russian data. Histograms are 5-year age-class growth averages. Thin lines are their smoothed one standard error confidence limits. Thick lines are linear or negative exponential functions fit through the age-class data. The upper panels show the RCS curves derived using all available (i.e. living –tree and subfossil) data.. The lower panels show RCS curves derived separately from the Russian, living and subfossil data.”
If you compare Figure 1 and Figure 2, you will notice a big discrepancy between the RW values shown in the 1-5 ageclass in the bottom panel (about 1.1 mm) and the values in the top panel of Figure 2 (about 0.6 mm — the scale is x100). I have specifically checked these calculations to reconcile this difference. Figure 3 shows a histogram of ring widths for trees in ageclass 1-5 years — the high value in Briffa et al, 1996 is inexplicable.

Figure 3. Polar Urals. Histogram of ring widths of “modern” trees aged 1-5 years. The value shown in NATO 1996 Figure 3 bottom panel is inexplicable.
Perhaps an even more serious problem occurs in the curve fitting. None of the parameters shown in Briffa et al. 1996 can be replicated from original data. Most problematic is the attempt to fit a negative exponential curve to the subfossil data. Here the problem is not simply parameter estimation: a negative exponential fit does not converge! If one re-examines the top panel of Figure 1, you will see that the curve is somewhat convex, which is why the negative exponential doesn’t fit.I’ve experimented with doing fits to all the data without previously taking 5-year agegroup averages and again did not obtain convergence for subfossil RW data (and different parameters again for other series).
The somewhat convex character of the subfossil data (especially when combined with an examination of original curves) suggests to me that there is a systemic problem in which early portions (“heartwood”) of subfossil samples are missing. “Heartrot” is a very common condition with conifers, and perhaps even particularly with larch. I’ve googled a bit on this. This link discusses fungi causing heartrot, which seems to occur more with dead trees than living trees.
I am not in a position to conclude whether the discrepancy between subfossil trees and living trees is due to a “heartrot bias” or to something else. However, the statistical differences are sufficient that one can firmly point to a lack of homogeneity between the modern and subfossil samples. The bias will make the subfossil cores date younger than they appear to actually be; under RCS methods, the RCS ring widths for subfossil periods will be lower than it appears that they should be. Additionally, what appear to be numerical problems in the Briffa parameter calculations will bias these RW calculations even more.
Now the Briffa temperature reconstruction for the Polar Urals is driven mostly by MXD rather than RW values. If the ring ages of the subfossil trees are underestimated by the year 1 assumption, then this will also affect the MXD calculations. Impressionistically, in the subfossil data, one sees a tailing off of MXD values in the last years of a tree’s life. This is a common pattern in the individual plots — the opposite of a juvenile growth pulse, if you will. The living trees do not show this dying “wilt”. The RCS model for MXD is linear: this will tend to allocate dying wilts in subfossil data to climatic effects, thereby biasing downward the calculations. I don’t know whether this effect is material, but it does seem observable.
The Tornetrask “adjustment” is driven by RW series; the "heartrot bias" may affect the Toronetrask "adjustment". I haven’t checked the impact of this effect on the Tornetrask data yet, but plan to do so.





One Comment
This is a different issue conceptually and at a different level of speculation and covering different territory (general methodoloty versus specific study). no reason to link it with Polar Urals. You link it because you’re concentrating on “mistakes by stick team” or even “bias by the stick team” rather than “what is interesting out there in the science”. but from a science standpoint, they are seperate methodological issues. Different pubs, Steve.
From a methodological standpoint, I have a lot of questions about the RCS. Implicitly they are trying to do the right thing by decomvoluting a confounding variable, but I still wonder about things like survivorship bias, epoch differences, etc. Would be easier to tear apart the RCS if it were a more established technique. Maybe you should even suggest the right way (best way, least worst way) to do RCS, Steve.